Scientific Name
Chilabothrus strigilatus fosteri

The subspecies was described and named by Dr. Thomas Barbour (1884-1946), a zoologist at the Harvard Museum. Barbour was, at one time, the Director of the Harvard Biological Station and Botanical Garden of Soledad, Cuba.
Holotype
MCZ no. 46054 from near Alicetown, North Bimini, Bahamas. Collected by Richard W. Foster and John H. Huntington in June, 1941. There are six paratypes, no’s 46055-46059, 46178; all from near Alicetown, North Bimini, Bahamas.
Type Locality
North Bimini, Bahama Islands.
Synonyms
- Epicrates striatus fosteri : 64 [65] [61] [62] [63]
- Epicrates striatus fosteri
- Epicrates striatus fosteri : 259
- Epicrates striatus fosteri
- Epicrates striatus fosteri [86] [87]
- Epicrates striatus fosteri
- Epicrates striatus fosteri
- Epicrates striatus fosteri : 56
- Epicrates striatus fosteri
- Epicrates striatus fosteri
- Epicrates striatus fosteri : 21
- Epicrates striatus fosteri Tipton, 2005: 46
- Epicrates striatus fosteri : 98
- Chilabothrus strigilatus cf. fosteri
- Chilabothrus strigilatus fosteri
- Chilabothrus strigilatus fosteri : 15
- Chilabothrus strigilatus fosteri : 282
- Chilabothrus strigilatus fosteri : 38
Barbour named C.s. fosteri after Richard Winslow Foster, who led a team to investigate the fauna of the Bimini Islands. The team returned with seven boa specimens, all from North Bimini.
Common Name
Bimini Boa.
Description and taxonomic notes
Barbour (1941) described and named Epicrates (then) striatus fosteri using scale counts provided by Arthur Loveridge and compared the data with Stull’s borrowed manuscript. Barbour determined there were no differences other than coloration that separated the Bimini boa from any of the other striatus . Based upon Barbour’s description Sheplan and Schwartz state, “no other Bahamian Epicrates has been so poorly diagnosed.”

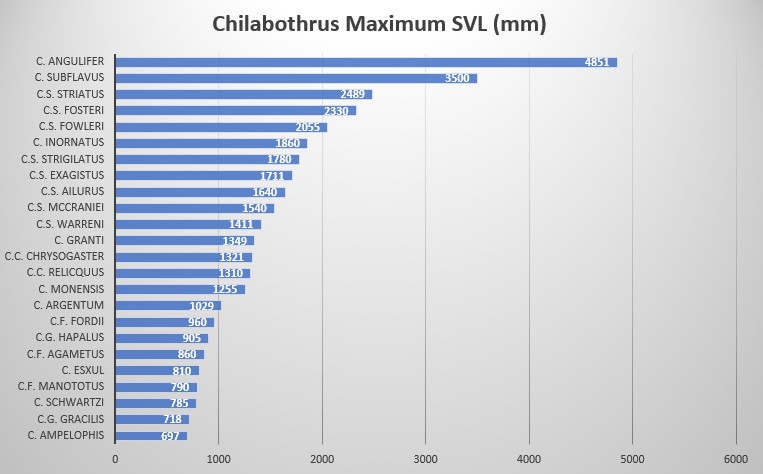
Chilabothrus strigilatus fosteri is characterized by its large size: 2330 mm for males and 1895 mm for females, 272-286 ventrals for males and 269-282 for females, 1 intersupraocular scale (modally), 14 supralabials with 3 scales (7-9) entering the eye, 10 scales in the circumorbital series (modally), 1 loreal (modally). The body and tail blotching cannot be counted because of the fusions between them; the interspace counts on the body generally are between 66 to 108 and 4 to 16 on the tail.

C.s. fosteri differs from all other subspecies (sans mccraniei) by its very dark dorsal coloration. The “black as a ravens wing” used by Barbour (1941) is rarely found; the more common colors of very dark brown to very dark gray are more the norm for the subspecies. None of the other striatus from Hispaniola or east of the Tongue of the Ocean come close to this character in fosteri. C.s. fosteri differs from C.s. mccraniei in that there is:
- complete fusion of the dorsal blotching vs. complete separation in mccraniei.
- one vs. two supraoculars.
- higher ventral and ventral + subcaudal counts.
- it differs from all other Hispaniolan and Bahama populations in that it has three supralabials entering the eye (7-9).
- all other subspecies have two supralabials entering the eye (7-8 and 8-9 modally).
A recent molecular phylogeny came to conclude that the two subspecies C. s. fosteri and C. s. fowleri form a single clade. It was therefore suggested to treat C. s. fosteri and C. s. fowleri as a single form termed Chilabothrus strigilatus cf. fosteri . While the authors did a thorough analysis and used only specimens with confirmed origin, it should also be noted, that the sample size for the study was overall relatively small and in case of the Bimini and Berry Island boas consisted of a total of four specimens (1 Bimini Boa, 3 Berry Island Boas). While this analysis delivered results consistent in itself, we are cautious to follow their argumentation for several reasons and thus continue to treat the currently described subspecies of C. strigilatus as such. The traditional view of the subspecies was recently employed by the lead author of the study in later papers . Thus, until further research provides strong evidence to support the 2013 viewpoint, we consider all subspecies described so far as valid.
C. s. fosteri is the subspecies occurring northernmost in the distributional range of the species. The prominent dark coloration might be an adaptation to cooler climates on these northernmost islands. A similar chromatological-geographical difference is present in Morelia s. spilota and Morelia s. variegata in Australia. However, without further analysis it is impossible to differentiate between evolutionary advantage through coloration and a founder effect unlinked to evolutionary constraints. If indeed the coloration reflects an adaptation to cooler climate, C. s. fosteri must be considered an Evolutionary Significant Unit (ESU) which needs to be treated and protected differently from C. s. fowleri, even if the lineage divergence occurred relatively recently and would not, per se, justify a splitting into two subspecies.

Evolution
Neither fossil or subfossil remains of Chilabothrus strigilatus fosteri have been found to date. A study based on molecular data (while not discriminating between the different subspecies) suggests that Chilabothrus strigilatus is on an isolated evolutionary trajectory since 2.6 MYA .
Distribution
East Bimini, Easter Cay, North Bimini and South Bimini.
[intergeo id=”AM2ETM”][/intergeo]
The five subspecies of Chilabothrus strigilatus occur on different islands of the Bahamas. The flags on the map indicate a type locality of a Chilabothrus strigilatus subspecies. Click on each flag to see which subspecies occurs on that particular island.

* Source
Habitat
In the Bahamas the mean annual temperature is 24.73°C (76.51° F) and the mean annual precipitation is 1264.46 mm. The islands experience warm, humid conditions year-round, though with more seasonal variations than the Southern Caribbean islands. There are also variations between the islands of the Bahamas, with rainfall falling twice as much in the northwestern islands than in the southeastern islands, and the more northerly islands experiencing temperatures up to 5° cooler than the southern islands.
Average temperatures are fairly high, with the mean daily temperatures fluctuating between 17°C and 32°C (62.6° F and 89.6° F). Mean annual rainfall for the Bahamas varies from about 865 mm to about 1470 mm. Inter-annual variability in climate is strongly influenced by the El Niño Southern Oscillation (ENSO). El Niño episodes bring warmer and drier conditions between June and August. Located in the heart of the Atlantic hurricane belt, the Bahamas is also subject to hurricanes and tropical cyclones especially during the August – November period.
Mean temperatures have increased by around 0.5°C since 1960, at an average rate of 0.11°C per decade. Bahamian data show that the mean daily maximum temperature for July has increased at a rate of 2°C per 100 years, and more recently at a rate of 2.6°C per 100 years. There have been statistically significant increases in the frequency of ‘hot’ days and nights, and decreases in ‘cold’ days and nights during the period 1973-2008.
There is seasonal variation in the rate of temperature increase, with the rate being most rapid in the warmest seasons, June-August and September-November, having rates of 0.13 and 0.15°C per decade respectively. There is also variation between islands, where the rate of warming is more rapid in the northeastern islands compared to the southwestern islands.
C.s. fosteri are found mainly on East Bimini. North Bimini has been, for all intents and purposes, cleared of all vegetation except for the mangrove in the west. Invasive casuarinas are in much of the drier land. South Bimini has also been cleared of vegetation in the west. The wetlands have been mostly reclaimed for development .





Longevity
The only longevity record for Chilabothrus strigilatus fosteri lists a captivity record of 10 years and 11 months. The animal was a wild caught female of unknown age at capture, kept at the Fort Worth Zoo, Texas . Considering that the nominate subspecies is long lived (20+ years is the average-not the exception), we assume that C. s. fosteri can live equally as long.
Behavior
Chilabothrus strigilatus are very active boas; they are excellent climbers as noted in Henderson (XXXX) . A boa was found as high as five meters up in a tree. In Terraria, the boas are always curious observing the area outside their terraria carefully and take the opportunity to explore. They seem to be docile, yet when initially handled they appear a bit nervous, but calm down when finally in the hands of the keeper. They don’t tend to bite but rather flee when disturbed.
Diet
In the wild the young take small lizards (Anolis) as first foods, like the remainder of the genus (with the exception of C. angulifer). With growth the boas eventually shift their diet to rodents (introduced Mus and Rattus), birds, fowl and their eggs. A small C.s fosteri (ASFS V7121) contained a catbird (Dumetella) .
In captivity the neonates start on ectothermic prey such as Anolis sagrei. It is possible that one or two from a litter might take a live pink mouse but they are the exception to the rule. Once the babies are feeding regularly they can be switched to pm by scenting them with anole skin, blood from the tail of an anole or chick fuzz. At this point the growing boas take appropriately sized rodents (rats and mice), chicks and quail.
Reproductive biology
Slavens reports the following reproduction data : C.s. fosteri: 1979 METF 0.0.2 born during 1979.
1980 JACF 0.0.8 born during 1980. The International Zoo yearbook lists for 1978 two breedings of C. s. fosteri : one at the Reptile Breeding Foundation in Picton, Canada, which produced 15 young and the other at the Jacksonville Zoo and Gardens, producing two young . The Jacksonville Zoo and Gardens might have been one of the first institutions to succeed in captive breeding of the Bimini Boa; their first litter was produced in 1972 .
Reproduction is triggered, according to Tolson and Huff, by regularly misting the enclosure during the months March through May. In captivity the boas mate in the months March through May and parturition occurs in the months September through November. In captivity multiple males have been observed breeding a single female. This behavior is also exhibited in mating C. inornatus. Neonate SVL is 318 mm to 500 mm .

Captive management
See Chilabothrus strigilatus account.
Conservation status, threats and population size in nature
CITES: Appendix II
Bahamas joined CITES on 18 September, 1976
IUCN Red List: Least Concern (LC)
Catalogue of Life: (click here)
The National Center for Biotechnology Information: (click here)
CITES import/export data: (click here)
Thomas Barbour (1941) writes: “This sparsely settled and, for years, utterly unimportant little archipelago has recently become famous as a fishing ground. Luxurious camps transformed some of the cays while others have been bought privately. This has resulted in the extermination of what few land vertebrates were once found on such islands as Cat Cay and Gun Cay. There is a single, longstanding native settlement on North Bimini, Alicetown by name; and a few scattered farms and a house or two on South Bimini, which is otherwise uninhabited.”
Sheplan and Schwartz (1974), while re-describing C. s. fosteri state the boa is “common on all four islands.” Since then, the boas numbers have declined as a result of development, road mortality and the international trade (which has not been a factor since 2000-see CITES export data above). How times have changed for this exquisitely unique boa. C. fosteri was listed in 1990 as a rare (R) species by the IUCN. A listing of (R) was defined as “taxa with small world populations that are not at present “Endangered” or “Vulnerable”, but are at risk .

At the time of this writing (Sept. 2019) IUCN lists the conservation status of this species as least concern (LC), however, based on the observation that the number of mature individuals is decreasing they conclude that the population is decreasing as a whole. Unfortunately a lack of data makes it impossible to say with certainty how steep the decline is, if any, and how large the population total is. As for all Island fauna, the restricted range they inhabit is a constant threat for species survival.
We disagree with the IUCN rating of LC and consider the Bimini Boa one of two (C. exsul being the other) most endangered boas in the West Indies. Development of North, East and South Bimini has destroyed almost all the habitat for these boas. Road kills on all 3 islands add to the mortality rates; adult boas are a very rare sight as a result of the roads (Crutchfield pers. comm.). The plight of the Bimini Boas cannot be overstated. At the current rate of development and road mortality rates, this beautiful and unique boa may be nothing more than a photo on this website and elsewhere in the near future.
Data taken from Buckner et al. was used to generate the table below:
Taxon |
Number of islands |
combined total surface area in km2 |
| Chilabothrus s. strigilatus | 9 | 1548.57 |
| Chilabothrus s. ailurus | 2 | 390 |
| Chilabothrus s. fosteri | 4 | 17.86 |
| Chilabothrus s. fowleri | 4 | 6211.75 |
| Chilabothrus s. mccraniei | 2 | 4.42 |
The total surface area of the islands as outlined here should not be falsely interpreted as suitable habitat. Detailed analyses about population size and suitable habitats are missing. We list this table to visualize potential threats as we consider it astonishing and worrying at the same time, that some subspecies formed on very small islands. It is worrying that the cane toad Rhinella marina has been recorded from New Providence as well as from Abaco Islands in the West Indies . Considering the devastating impact it had on the Australian herpetofauna, we hope that the West Indian species are less impacted by its occurrence.
The CIA World Factbook lists the following environmental threats for The Bahamas: coral reef decay; solid waste disposal .
The maps below illustrate the extent of habitat destruction and alteration due to development and agriculture.


To add an historic note to the map from 1737 is a passage from Catesby (1743) regarding the Bahamas iguana population and its role in the trade and food chain. He writes, “These Guana’s are a great Part of the Subsistance of the Inhabitants of the Bahama Islands, for which Purpose they visit many of the remote Kays and Islands in their Sloops to catch them, which they do by Dogs trained up for that Purpose, which are so dexterous as not often to kill them, which if they do, they serve only for present spending; if otherwise they sew up their Mouths to prevent their biting, and put them into the Hold of their Sloop till they have catched a sufficient Number, which they either carry alive for Sale to Carolina, or Salt and barrel up for the Use of their Families at Home. These Guana’s feed wholly on Vegetables and Fruit, particularly on a Kind of Fungus, growing at the Roots of Trees, and of this and others of the Anoma Kind.”
Population in captivity
We are aware of no known specimens in public institutions; there are 5 known adult specimens in the private sector in the US. To date, all reproduction efforts have failed to produce any viable young. The subspecies has been kept in the EU in the 1980s-90s but we have no information of any specimens currently alive in Europe. The Bimini Boa should be a priority for both Public Institutions and the Invisible Ark.
On display in these Zoos
We are unaware that this species is kept in any zoological institution worldwide.
Beastly Biminians


 near the Bimini Sands Resort on the coast of South Bimini. Photo Emily Powell")













Several Pictures of Chilabothrus strigilatus fosteri from various sources (link outs)
Chilabothrus strigilatus fosteri from South Bimini, Bahamas
Chilabothrus strigilatus fosteri from South Bimini, Bahamas

Citations