Scientific Name
Chilabothrus fordii agametus

The subspecies was described and named by Sheplan & Schwartz. Bruce R. Sheplan was a herpetologist at Miami-Dade Community College. Albert Schwartz (1923 – 1992) was a Professor of Biology at Miami-Dade Community College. Both co-authored the seminal work "Hispaniolan boas of the genus Epicrates (Serpentes, Boidae) and their Antillean relationships."
Holotype
MCZ no. 62656. Adult male from Mole St. Nicholas, Department du Nord-Ouest, Haiti. Collected by James D. Lazell and A. Stanley Rand on 26 July, 1960.
Type Locality
Môle St.-Nicolas, Département du Nord-Ouest, Haiti.
Synonyms
-
- Epicrates fordii agametus [111] [112]
- Epicrates fordii agametus
- Epicrates fordi agametus
- Epicrates fordi agametus : 45
- Epicrates fordi agametus Henderson, Schwartz & Inchaustegui, 1984: 93
- Epicrates fordii agametus
- Epicrates fordii agametus
- Epicrates fordi agametus : 19
- Epicrates fordii agametus Crothers, Powell, et al, 1999: 114
- Epicrates fordii agametus
- Epicrates fordi agametus Tipton, 2005: 45
- Epicrates fordii agametus : 132
- Chilabothrus fordii agametus : 11
- Chilabothrus fordii agametus : 38
The name agametus is derived from the Greek for "bachelor" referring to the unique male holotype .
Common Name
Môle St.Nicolas boa.
Description and taxonomic notes
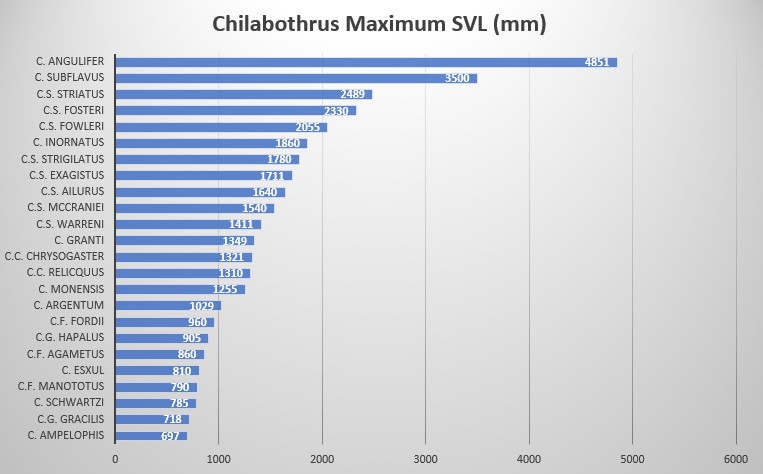
It is assumed Chilabothrus fordii agametus is sexually dimorphic. This subspecies is described from an adult male with a SVL of 860 mm, tail 170 mm, ventral scale count of 261, subcaudal scales are 82 (unpaired), ventrals + subcaudals are 343, anal scale entire, dorsal scales at neck are 33, dorsal scales at midbody are 38, anterior to vent 22 scales, supralabial scales are 13-14, supralabials entering the eye are 6-7 and 7-8, infralabials are 15-16, loreals are 2-2, preoculars are 2-2, postoculars are 6-7, circumorbitals are 10 & 13, intersupraocular scale is 1 bordered by 3 scales anteriorly and 5 scales posteriorly.
C.f. agametus has 92 dark dorsal body blotches and 17 dark dorsal tail blotches . C.f. agametus and C.f. manototus are easily differentiated by the number of dorsal blotches; 92 in C.f. agametus and 72-74 in C.f. manototus. While the the difference in blotching is relatively small between C.f. agametus and C.f. fordii, there is a possibility C.f. agametus is the larger of the two, based on the long series of specimens examined .
Chilabothrus fordii is one of the smallest species within the genus. Neonate C.fordii (and C. monensis) are the only members of the genus born with a gray or grayish-brown ground color .

*Source
The two subspecies Chilabothrus fordii agametus and C. f. manototus were described by Sheplan and Schwartz (1974) and Schwartz (1979) on the basis of one C. f. agametus and two C. f. manototus specimens. While a small sample size should in general not hinder the description of a species or subspecies, it is crucial to remark at this point, that some doubt exists over the justification of both subspecies.




Schwartz writes in 1979, while describing C.f. manototus :
There are also apparently isolated populations at Cap-Haitien on the northern Haitian coast and in the Valle de Cibao in northwestern Republica Dominica; these 2 populations are questionably assigned to E.f. fordii (Sheplan and Schwartz). A third isolate has recently been discovered on Isla Catalina by R.I. Crombie (pers. comm.). That island is far (135 km) to the east of the previously easternmost locality record (Limonal) for the species.
Quite interesting is the following remark by Schwartz and Henderson :
"Epicrates fordi is known also from the northern Dominican Valle de Cibao (between Monte Cristi and Villa Vasquez); the population remains unassigned sub-specifically".
The current situation makes it difficult to justify the status of the subspecies C.f. agametus and C.f. manototus. We recommend examining both subspecies, morphologically and genetically, to determine their validity as subspecies. This should include specimens from across Hispaniola to clarify the subspecific questions and any possible intergrading.
Distribution
The full distribution of the subspecies has not been entirely determined. It possibly occurs throughout the Presqu’ile du Nord-Oest; otherwise known only from the type location.. The taxonomic uncertainty lasting over the three C. fordii subspecies makes it difficult to determine if intergradation zones exist, and if so, where?
[intergeo id=”ANzETM”][/intergeo]
Three subspecies of Chilabothrus fordii occur on Hispaniola. The Flags on the map indicate a type locality of each subspecies. Scroll over to see which subspecies occurs at the indicated point.
Habitat
Hispaniola consists of Haiti (the “hilly country”) and the Dominican Republic. There are four main mountain ranges on the Dominican Republic side that parallel each other while running northwest to southeast. Cordillera Septentrional is the most northern range. Cordillera Central has the highest point on the island, Pico Duarte at 3087 meters, and extends into Haiti. The two ranges in the south, Sierra de Bahoruco and Sierra de Neiba, rise in excess of 2000 meters. A smaller range, Cordillera Oriental, rises to 600 meters and is also in the northeast. A majority of the island is over 1000 meters in elevation.
Haiti also has its share of mountain range chains. In the southwest is the Massif de la Hotte and Massif de la Selle in the southeast. The Massif de la Selle , with Pic la Selle that rises to 2674 meters, runs eastward to become the Sierra de Bahoruco. In the central area are the Montagnes du Trou-d’Eau and the Chaine des Matheux. In the north is the Massif du Nord and in the north center is the Montagnes Noires. In the northwest is the peninsula, Presqu’ile du Nord-Oest, a low ridge with arid areas.
The mean annual temperature is 24.40°C and the mean annual precipitation is 1438.40 mm. Located in the Caribbean’s Great Antilles, Haiti has a hot and humid tropical climate. Daily temperatures typically range between 19°C and 28°C in the winter and 23°C to 33°C during the summer months. Northern and windward slopes in the mountainous regions receive up to three times more precipitation than the leeward side. Annual precipitation in the mountains averages 1,200 mm, while the annual precipitation in the lowlands is as low as 550 mm. The Plaine du Gonaïves and the eastern part of the Plaine du Cul-de-Sac are the driest regions in the country. The wet season is long, particularly in the northern and southern regions of the island, with two pronounced peaks occurring between March and November.
Mean temperatures have increased by 0.45°C since 1960, with warming most rapid in the warmest season, June-November. The frequency of hot days and hot nights increased by 63 and 48 days per year, respectively, between 1960 and 2003. The frequency of cold days and cold nights has decreased steadily since 1960. Mean annual rainfall has decreased by 5 mm per month per decade since 1960. The intensity of Atlantic hurricanes has increased substantially since 1980.
Chilabothrus fordii agametus: Based on Lazell’s (1961) description of the Mole St. Nicholas the area is “arid,” and he implied that Acacia is common there. This is the sort of ecological situation where C. fordii occurs most abundantly elsewhere .

Longevity
The nominate species, Chilabothrus fordii, seem to live, in general, quite long; there is no reason to believe the subspecies are less so. There were no live specimens from which to gather data from.
Reproduction
Based on the weather, geography and proximity to the nominate species, it would not be unreasonable to attribute the same mating behavior, gestation and parturition data to the subspecies. Locality differences in climate and habitat may influence the start and end period for mating and parturition. No data regarding the reproductive biology of this subspecies exists. We assume this taxon has a similar reproductive biology as the nominate subspecies.
Behavior
In lack of any studies or anecdotes about the behavior of C. fordii agamteus we can’t say with certainty if this subspecies differs from the nominate ssp. However we strongly assume that the behavior of the Môle St.-Nicolas Boa is very similar to its closest relative, the Ford’s boa. Most likely a crepuscular to nocturnally active boa, capable of secreting a strong (and not pleasant) smelling musk. Sheplan and Schwartz stated that they have never been to the type locality . They refer to Lazell’s description of the area as arid and implied that Acacia is common . This description reminded Sheplan and Schwartz of the habitat type which is favoured by C. f. fordii and thus believe that the ecology of C. f. fordii and C. f. agametus is similar.
Diet
Like the nominate species, it can be assumed the subspecies’ diet in the wild consists of Anolis and small rodents (Mus) . Henderson et al. examined the stomach contents of 48 C. fordii. The boa was found to prey upon Anolis cybotes, Anolis sp. and small rodents (Mus).
Captive management and captive population
We are unaware that this subspecies has ever been kept in captivity.
Conservation status, threats and population size in nature
Haiti is not a member of CITES.
Dominican Republic joined CITES on 17 December, 1986; entry into force on 17 March, 1987.
IUCN Red List: None
Catalogue of Life: (click here)
The National Center for Biotechnology Information: (click here)
CITES import/export data: (click here)
Nellis et al. reported in 1983 that Chilabothrus fordii occurs at high population density in some areas and question the dietary reasons for its success”. . 17 years later, Powell and Co-workers report that all three Dominican Boid species (Chilabothrus fordii, C. gracilis, C. striatus) “appear to be sustaining viable populations, sometimes even in heavily disturbed areas”. Despite this finding, they consider the protection status for the three species justified, because of their vulnerability to exploitation by the pet trade and, to a lesser degree, their dependence on threatened habitats . We would reverse these concerns as there is no pet trade regarding any of the West Indian Boas. These boas were last imported in 2003 in minute numbers. See the CITES data above.
The last import of boas from Hispaniola was in 2003; the pet trade is no longer a threat, and has not been for seventeen years. It should not be a consideration of any consequence when determining population numbers or designing conservation measures. These snakes remain abundant in some areas, and the CITES listing reflects presumed threats emanating from international trade for the pet industry, which currently are not applicable to these species.
Powell and Incháustegui, two of the authors from the above study, report in a study published in 2009 that the introduced species, Bufo marinus has become widely distributed and is invading even very xeric areas that provide breeding sites only intermittently . While their study did not conclude to what extent Bufo marinus is affecting the Hispaniolan ophidian fauna, it can be estimated from the situation in Australia, where Bufo marinus was introduced as a Neozoon , that it will have a significant influence on population and conservation status of different snake species on the island of Hispaniola.
It should be also taken into consideration that the situation on Hispaniola, especially Haiti, has worsened for wildlife in general. The 2010 earthquake left Haiti largely ungoverned and the humanitarian crisis was followed by an ecological crisis due to destruction of habitats. Thus making it difficult to envisage a stable future for the snake species on the island. In addition to this, climate change is a factor which is likely to gain momentum in ecosystem changes throughout the West Indies. Giant storms fueled by rising oceanic temperatures as a consequence of human caused climate change are more likely to form more often than before. On the other hand, climate change will also increase droughts and diminish fresh water resources on the islands of the West Indies.
In a relatively recent study by Hedges et al. they predict, through modeling, that Haiti will experience mass extinction events by 2036 as a result of current deforestation trajectories. They comment: “We project that all primary forest in Haiti will disappear by ∼2035 (CI, 2033.5–2035.4) at the current rate, defining phase II as a 49-year period from 1986 to 2035 during which the final 4.8% of primary forest in the country will be lost. Assuming that our estimated loss of vertebrates is representative of the biodiversity in general, then 66–83% of species will be lost in Haiti during 1986–2035 because of deforestation. Thus, Haiti is well into a mass extinction, with only 8 of 50 mountains still holding primary forest. Unfortunately, Haiti’s neighbor, the Dominican Republic, is not a major refuge for Haitian species because more than half of the species surveyed (51%, average) on mountains with primary forest are endemic to Haiti and 12% are endemic to an individual mountain in Haiti. In addition, forest loss in the Dominican Republic is a threat to that country’s biodiversity.”
They continue: “It is common for deforestation to occur within protected areas of tropical countries, and, during the course of our surveys in Haiti, we observed ongoing destruction of primary forest inside all of the national parks. We also estimate that 60–75% of primary forest in the two original national parks, Pic Macaya and La Visite, has disappeared since they were declared as protected areas 35 years ago. In both cases, the rates of primary forest loss (pre-2000 and post-2000) were greater than the overall rates for all of Haiti, indicating that protection was minimal or nonexistent. This indicates that the mass extinction of biodiversity in Haiti will continue unabated unless greater protective measures are taken. More generally, this suggests that the phrase “protected area” be reserved only for areas where protection has been confirmed.” , .
A study on the risks for species extinction on island ecosystems worldwide analysed 15 insular regions, applying novel network analysis methods which enabled to disentangle the associations of multiple threats on vertebrates, invertebrates, and plants. The results showed that Biological invasions, wildlife exploitation, and cultivation, either alone or in association, are the three most important drivers of species extinction and decline on islands to date . These results are consistent with expectations when analysing past and even current events. However, considering that climate change will dramatically change whole ecosystems in previously unprecedented ways, we estimate that this threat is more likely to affect species survival on an overall scale than the past drivers of extinction.
For a nation struggling to feed and house its’ people, conserving its fauna and herpetofauna are far from the top of the list, if they are on the list at all. Granted, there are measures in place, all on paper, that lay aside land for National Parks. Yet enforcing the measurements laid out on paper are anything but enforceable, as noted above. It is truly a travesty of the highest order; we can only hope some of the herpetofauna escapes the sickle of extinction.
Thus we consider the biggest threads for species survival in (1) large scale ecosystem changes, (2) invasive species, (3) climate change.
We feel, given the boas’ extremely small ranges, few museum specimens of C.f. agametus & C.f. manototus, and the current rate of habitat destruction taking place, it is imperative to determine the current status of the three subspecies as quickly as time and resources allow.
The CIA World Factbook lists the following environmental threats for Haiti: extensive deforestation (much of the remaining forested land is being cleared for agriculture and used as fuel); soil erosion; overpopulation leads to inadequate supplies of potable water and and a lack of sanitation; natural disasters .
The map below is to illustrate the extent of habitat destruction and alteration due to development, intentional deforestation and agriculture.


On display in these Zoos
We are unaware of any live Chilabothrus fordii agametus in Zoo or Academic collections anywhere in the world. If you have further information, please Contact us.
